lunes, 27 de octubre de 2014
martes, 21 de octubre de 2014
martes, 7 de octubre de 2014
domingo, 5 de octubre de 2014
Lectura / Hormonas - 904
Los orígenes evolutivos de las hormonas, neurotransmisores, y otros mensajeros químicos extracelulares
Seminarios de medicina del Hospital Beth Israel, Boston
- 11.06.2000 -
Las implicaciones para la biología de mamíferos
Jesé Roth M.D., Derek LeRoith M.D., Ph.D, Joseph Shiloach, Ph.D., James L. Rosenzweig, M.D., Maxine A. Lesniak, B.S., and Jana Havrankova, M.D.
En años recientes, ha habido observaciones de que algunas hormonas peptídicas de los vertebrados están presentes en invertebrados multicelulares – no solo en aquellos complejos como las moscas, sino también en las formas multicelulares más simples.
En este trabajo resumimos la evidencia que dice que hasta los organismos simples unicelulares producen moléculas que son muy similares a las hormonas peptídicas de los vertebrados. Además, existe evidencia de que los elementos bioquímicos que asociamos con la acción hormonal también están presentes en organismos unicelulares, aun en la ausencia de las unidades anatómicas que asociamos con los sistemas endocrino o neuroendocrino, como las glándulas, nervios, el sistema circulatorio, y células blanco diferenciadas.
Equipados con la información de que las hormonas y los elementos bioquímicos relacionados probablemente se originaron con los organismos unicelulares, reexaminamos un gran numero de fenómenos en los seres humanos y otros vertebrados. Esto explica muchas cosas, incluyendo la superposición del sistema endocrino con el sistema nervioso, la superposición de las hormonas con los factores tisulares, la producción normal y ectópica de hormonas y los mecanismos endocrinos y exocrinos.
Las Hormonas en Organismos Unicelulares
Se detectó material similar a la insulina en algunas especies de protozoarios, dos especies de hongos, y tres cepas de Escherichia coli. Además se detectó material similar a la somatostatina, ACTH, y β-endorfina en protozoarios. Parece que estos materiales de tipo hormonal son muy similares a sus contrapartes de vertebrados, especialmente en la conservación de las regiones tridimensionales que son necesarias para el reconocimiento por receptores específicos de los vertebrados y por anticuerpos específicos.
Debemos enfatizar que la primera demostración clara de la presencia de un péptido hormonal de vertebrado en un organismo unicelular fue la detección de un material similar a la gonadotrofina coriónica humana en extractos bacterianos.
Otras moléculas mensajeras de vertebrados han sido encontradas en organismos unicelulares. Estas incluyen neurotransmisores (catecolaminas, serotonina, y acetilcolina) y esteroides que tienen muchas similitudes con sus contrapartes en vertebrados. (En nuestro trabajo nos hemos concentrado en los mensajeros peptídicos porque su presencia requiere que el organismo halla heredado y mantenido el correspondiente ADN; en el caso de otros tipos de mensajeros, como las aminas y esteroides, que son fácilmente derivados de constituyentes celulares comunes, podría ser discutido si su presencia es azarosa.)
Evidencia de Receptores Hormonales y Efectores en Organismos Unicelulares
Se ha observado que muchas hormonas de vertebrados tienen efectos biológicos en los organismos unicelulares, pero solo en dos casos se sabe que están mediados por receptores específicos: la epinefrina estimula la adenilato ciclasa en protozoarios –un efecto que es bloqueado por un receptor específico antagonista, el propranolol; y los péptidos opioides y alcaloides alteran la conducta de alimentación de las ameabas –un efecto que es bloqueado por naloxona, un inhibidor especifico a nivel del receptor.
Aunque las moléculas de tipo hormonal y los sistemas receptor-efector de los vertebrados han sido detectados en organismos unicelulares, se debe enfatizar que no se ha demostrado su función dentro del organismo unicelular. Sin embargo, los sistemas intercelulares humorales de comunicación que regulan la reproducción sexual y las reacciones hacia la comida han sido descriptos en organismos unicelulares. Por ejemplo, en levaduras y otros microbios eucariontes, los péptidos y glicoproteinas han sido identificados como moléculas mensajeras.
Resumiendo: esta información parece sugerir que los elementos bioquímicos que son usados por el sistema endocrino, el sistema nervioso, y otros sistemas de comunicación intercelular de los vertebrados probablemente se originaron en organismos unicelulares. Con la evolución, los elementos anatómicos del sistema (la célula que produce el mensajero, la célula que responde a él y el fluido extracelular que lo transporta) se han hecho extremadamente complejos, especializados y diversos; por otro lado, los elementos bioquímicos se han conservado.
Fenómenos de Vertebrados Reexaminados
Superposición de los sistemas endócrino y nervioso
En los vertebrados, los dos sistemas dedicados a la coordinación son el sistema endócrino y nervioso. Tradicionalmente, estos sistemas eran vistos como entidades separadas, pero la línea que los separa ha sido penetrada de varias maneras.
Las neuronas aparecieron primero con los organismos multicelulares más simples, pero las glándulas endocrinas típicas aparecieron junto con los vertebrados más simples. Pearse y Polak y sus colaboradores han interpretado estos descubrimientos como indicación de que el sistema nervioso es filogenéticamente más viejo y dió orígen al sistema endócrino de los vertebrados. También han sugerido que embriológicamente el sistema nervioso antecede al sistema endócrino y que las neuronas de la cresta neuronal son los precursores embriológicos de las glándulas que producen las hormonas gastrointestinales.
De acuerdo a nuestra formulación, el sistema nervioso no origina al sistema endócrino, ni embriológicamente ni filogenéticamente. Lo que creemos que sucede es que ambos sistemas son derivados independientemente de un sistema ancestral común.
Agentes Parácrinos y Factores Tisulares
Muchas sustancias que parecen hormonas y neuropéptidos en su estructura y función no entran en el criterio de ninguna de las dos clases (insulin-like growth factors, tissue-specific growth factors, prostaglandinas e interferones). La incorporación de estos materiales dentro de los modelos de los sistemas endocrinos y nervioso ha sido difícil. Sugerimos que estos comunicadores intercelulares, incluyendo a las hormonas y neurotransmisores, comenzaron evolucionalmente y embriológicamente como factores tisulares locales actuando sobre las mismas células secretorias y sus vecinos; luego, con los avances morfológicos y de desarrollo, una parte de las moléculas mensajeras se convirtió en hormonas o neurotransmisores. Este punto de vista provee una racionalización para los descubrimientos de que muchas hormonas clásicas y neurotransmisores en mamíferos actúan igual fisiológicamente en el mismo organismo como agentes parácrinos o factores tisulares locales (somatostatina, insulina y glucagon). En esencia, la relación anatómica entre la célula secretoria y la célula blanco, al nivel del organismo multicelular, ha sufrido una evolución extensa y muestra gran diversidad. Por otro lado, la bioquímica fundamental del sistema por el cual un agente humoral transporta su mensaje desde la célula secretoria hasta la célula blanco es evolutivamente viejo y altamente conservado.
Superposición de las Funciones Exócrinas y Endócrinas
En los organismos unicelulares, la secreción de moléculas mensajeras es indistinguible de la secreción de las enzimas digestivas u otras secreciones que asociamos con los sistemas exócrinos de organismos complejos. En nuestro punto de vista, la creación de una diferencia entre el sistema exócrino y el sistema endócrino evolucionalmente necesitó de muchos rearreglos anatómicos que fueron adquiridos por los organismos multicelulares moderadamente complejos. Esto concuerda con el descubrimiento de que en vertebrados las células secretorias exócrinas y endócrinas son muy similares en apariencia, en la preparación de las proteínas para exportación y en los mecanismos de secreción. Un origen unitario de las funciones exocrina y endocrina también provee una explicación para el descubrimiento de que muchas moléculas mensajeras humorales, como el factor de crecimiento epidermal, factor de crecimiento neuronal, prostaglandinas, somatostatina, gastrina y prolactina están presentes (y funcionan) en fluidos exocrinos clásicos, como saliva, secreciones intestinales, leche y semen. De esta manera, un origen evolutivo común para los sistemas endocrino y exocrino provee la base para la superposición continua en los organismos complejos.
Conclusiones
El descubrimiento de que los organismos unicelulares producen péptidos hormonales ha sugerido varios puntos. Primero, todos los elementos bioquímicos de la comunicación intercelular surgieron muy temprano en la evolución y han sido altamente conservados. Segundo, el surgimiento de organismos anatómicamente complejos permitió la diferenciación y especialización de las células secretorias y células blanco, como también la diversidad de los compartimentos y fluido extracelular que transporta al mensajero difusible de célula a célula.
Finalmente, diversos fenómenos en los vertebrados pueden ser explicados ahora, incluyendo la superposición de los sistemas endocrino y nervioso, de hormonas y factores tisulares, de la producción normal y ectópica de hormonas y de los mecanismos exocrinos y endocrinos.
Tomado de: http://www.genaltruista.com/notas/00000177.htm
jueves, 25 de septiembre de 2014
Evolución de la unidad renal - 7°
Evolución
de la unidad renal
A medida que los vertebrados
fueron evolucionando se pudo observar como los órganos excretores se preparaban
para desempeñar su función excretora. En primera instancia se
puede observar que el glomérulo se comunican totalmente con el celoma del animal
de tal forma que el filtrado que se realiza no se puede atribuir en especial al
órgano excretor en sí, sino a una conjunto de interacciones entre celoma y
riñón (en esta instancia muy primitivo) el cual se denominó Holonefros
Probablemente el holonefridio consistió en dos series longitudinales de
unidades renales, formadas por un par de glomérulos en cada segmento
embrionario.
Estos órganos se pueden
mencionar como un vestigio de un riñón derecho y de un riñón izquierdo,
localización conocida como ratroperidional, es decir externa con relación con
el celoma o sometopleura. Los nefridios eran abiertos pues el túbulo estaría
provisto de nefrostoma, que todavía se comunicaba con la cavidad celómica.
Todos los túbulos desembocaban en dos conductos denominados arquinefridios, y
estos llegarían independientemente al exterior.
De estos en animales como
amocetos de mixinoides y anfibios apodos, los cuales presentan un solo par de
glomérulos por segmento. En cambio encontramos en estado embrionario el
denominado PROTONEFROS, en donde aparecen primero las unidades posteriores y
asumen desde luego su función de excreción. Existe pues una etapa ontogenia en
la cual el organismo está provisto de órganos excretores de posición muy
delantera, provistos únicamente de un par de nefridios por segmento, sin
importar que estos sean abiertos o cerrados, además los conductos urinarios son
del tipo arquenefridio. Los protonefros son
funcionales en amocetos, en las larvas de los teleostomos y aún en el adulto de
los ciclostomos. Los túbulos de protonefros descargan su contenido en un vaso
venoso. Opistonefros Con este nombre se designa el aparato renal que
representa, en los organismos adultos actuales, una porción posterior del
holonefros hipotético.
Aludiendo una vez más a la desaparición
de los elementos renales anteriores, diremos que esta prosigue hasta una zona
posterior al límite trasero de riñón de cabecera. Así, en los animales que
conservan tales restos de pronefros, queda un espacio intermedio, que no ejerce
funciones renales entre el riñón de cabecera y el opistonefros funcional. La generalidad de los
opistonefros se caracteriza por la multiplicación , que llegan a sobre pasar
excesivamente en número de segmentos correspondientes, sobre todo en la región
posterior que por tal motivo, aparece hipertrofiada ; además, en el
opistonefros, es notable la perdida de la organización segmentaria y carencia
casi universal de nefrostomas. Constituyen la excepción, los opistonefros de
los ciclóstomos que conservan la estructura segmentaria y tienen solo un par de
unidades por segmento.
De lo antes dicho, es
posible deducir que la diferencia fundamental entre el pronefros y el
opistonefros radica en la procedencia, a partir del holonefros, además de las
particularidades ya mencionadas, representa la porción anterior del holonefros,
es funcional en los embriones y en algunos estados larvario; si persiste es a
manera de riñón de cabecera. El opistonefros es manifestación de la porción
posterior del holonefros, es funcional en todos los peces, en el más amplio
sentido del término, así como en los anfibios aun cuando algunos animales
presentan características estructurales primitivas. Metanefros En párrafos
anteriores se hablo acerca de la hipertrofia de seccionposterior del
opistonefros y de la multiplicación de los nefrones en la porción
opistonefrica. Probablemente el fenómeno manifiesta cierta tendencia hacia la
constitución de los riñones a expensas de la parte más trasera del holonefros
hipotético. La tendencia llega a su culminación en los órganos renales de los
vertebrados amniotas, que son notablemente compactos y tienen enorme cantidad
de nefrones del tipo más complejo, antes descrito.
A esos órganos se les conoce
en el campo de la anatomía comparada con el nombre de metalíferos. Una vez más,
señalamos que aparte de las diferencias anatómicas, la peculiaridad básica de los
metalíferos es su origen a partir de la sección posterior del holonefros. Los
ciclostomos en general, están provistos de riñón de cabecera, constituidos por
residuos del pronefros embrionario, drena la cavidad pericárdica directamente y
en segundo término la abdominal, ya que ambas están en comunicación. En la
porción anterior de la zona pronefrica, se halla un espacio carente de unidades
renales y a continuación, se localiza el opistonefros con las características
antes descritas. Estos órganos desembocan por medio de dos conductos
arquinefricos simples, cada uno de estos se prolongan desde las primeras
unidades opistonefricas hasta le región cloacal, siguiendo el borde externo de
cada órgano urinario; cabe añadir, que por delante del punto inicial quedan
vestigios del conducto empleado por el pronefros embrionario o del estado
larval.
Poco antes de llegar al
exterior , ambos conductos que podemos llamar ureteros o conductos dewolff, se
unen en una sola vía que desemboca en el ápice de una papila pequeña, oculta en
la región posanal, dentro de una cripta poco perceptible. Solamente en los
machos adultos, durante la madurez sexual, la papila se hipertrofia y adquiere aspecto
de pene rudimentario.
Los elasmobranquios tienen
opistonefros típico, por lo general más voluminoso en la región posterior. En
estos animales, especialmente en el sexo masculino, son muy interesantes las
relaciones establecidas entre el aparato excretor y el genital. La disposición
y conformación del aparato excretor en los peces teleostomos, varían en los
diversos grupos; sin embargo, pueden indicarse algunas características
generales, como es la independencia en cuanto a la expulsión de los productos
genitales y urinarios, ya que el conducto arquinefrico es siempre funcional en
toda su extensión y los órganos reproductores generan nuevas vías de expulsión.
Desde luego que estos animales están dotados de opistonefros y en ciertos casos
los estados larvarios tienen restos de pronefros.
En los anfibios, la parte
anterior del opistonefro está relacionada con el aparato genital de los machos,
además, hay conductos accesorios que recogen que recogen la excreción de la
región posterior del opistonefros, pero en lugar de desembocar en la cloaca, se
unen a los conductos arquinefricos que son a la vez diferentes, y estos
descargan separadamente en la cloaca. En las hembras de estos
anfibios no existe relación entre el aparato genital y la porción delantera del
opistonefros, ni hay unión entre el conducto arquinefrico y el oviducto; el uno
y el otro son independientes y descargan separadamente en la cloaca. La
concentración de los órganos renales es más notable en los anfibios anuros; se
trata de un par de cuerpos ovoides, relativamente grandes y rojizos, a lo largo
de la cara ventral en cada riñón es perceptible una banda longitudinal
amarilla, la glándula adrenal.
Son pocos los túbulos que se
convierten en aferentes; sin embargo, el conducto arquinefrico es la única vía
de descarga para los productos genitales masculinos en la cloaca. No existe
relación entre los ovarios y el aparato excretor femenino. En ambos sexos, la
cloaca forma un receso que funciona como vejiga urinaria; los uréteres no
desembocan a tal vejiga si no en sus proximidades, la orina pasa del luman
cloacal al receso antes mencionado. En este grupo de vertebrados
el aparato urinario de los adultos aparece en forma de metalíferos. Es mucho más
compacto que en las formas anteriores y está completamente separado del aparato
genital , puesto que el conducto arquinefrico funciona solo como vaso deferente
y los riñones descargan por uréteres mas o menos complicados, pero siempre
independientes del conducto primitivo.
En los reptiles, la forma de
los riñones es variada, puesto que la morfología de estos animales lo es
también. Por ejemplo en las serpientes y en los lacertillos apodos, uno de los
riñones es anterior con respecto a un homologo, pero alojados, como sucede en
toda clase zoológica, en la región posterior de los cavidad abdominal. Todos los reptiles tienen
los riñones lobulados, a menudo alargados, y en algunas especies funcionados
entre sí. También debido a lo largo del cuerpo, los uréteres de las serpientes
son muy prolongados; en cambio, los cocodrilos y las tortugas tienen uréteres
cortos. Estos últimos animales están previstos de vejiga urinaria, originada en
parte por la cloaca embrionaria y en parte por la alantoides.
En aves y mamíferos; los
riñones son aún más compactos, siguen lobados y están alojados en la región
pélvica. Las aves en general carecen de vejiga urinaria; solo los avestruces
tienen, y los productos de la excreción se eliminan por la cloaca. El proceso evolutivo hacia
un estado más compacto llega a su máxima expresión actual en los riñones de los
mamíferos. Se trata de órganos cuya forma semeja un frijol, por tanto tiene un
borde convexo, que es el externo y el opuesto o interno, con una depresión
llamada hilio, que determina el punto de entrada para la arteria renal y de
salida para la vena respectiva y el uréter, así como para el paso de nervios y
vasos linfáticos.
Cada riñón está cubierto por
una membrana fibrosa, llamada cápsula renal, que en el hilio se prolonga sobre
la superficie del uréter. Si se observa un corte longitudinal del riñón humano,
encontraremos que hacia adentro de la cápsula existe un arrea compacta conocida
como corteza renal y más hacia la superficie del riñón y el ápice dirigido en
sentido inverso. Cada uno de dichos conos es
una pirámide renal. La región interna, correspondiente al borde cóncavo del
órgano, carece de pirámides; está ocupado por un espacio denominado pelvis, que
puede interpretarse como gran dilatación del uréter. La pelvis se resuelve
radialmente en estructuras en forma de embudo más próximas a la pelvis se
denomina cálices mayores, y cálices menores las inmediatas a las pirámides.
Los nefrones se encuentran
en la corteza y en la médula. Los corpúsculos renales o de mapigio , de los
cuales se desprenden, como se vio, el túbulo excretor están incluidas en la
zona cortical, inmediatamente después de la cápsula, conocida como de Bowman,
se presenta, todavía en la misma zona, la porción contorneada próxima del
túbulo, que continua por un tramo recto o descendente y de menor calibre,
dirigido, en la médula hacia el ápice de una de las pirámides. No llega hasta
tal extremo sino, que en sus inmediaciones forma el asa de Helen, y retorna en
sentido opuesto por medio de otro tramo recto, que se denomina ascendente,
hasta llegar nuevamente a la corteza. Una vez en este lugar, se hace sinuoso en
la región contorneada distal para finalmente desembocar en un tubo colector al
que confluyen numerosos túbulos. De nuevo el tubo colector va hacia la pelvis,
se une a otros semejantes y vierte su contenido en tubos excretores abiertos en
la cápsula de la pirámide correspondiente.
La orina es recogida por
cálices menores, de ahí pasa a los mayores, a la pelvis renal y posteriormente
al uréter. Los uréteres, uno de cada riñón, desciende hacia la región pélvica,
en la que se aloja la vejiga urinaria. Esta última descarga por un solo
conducto, la uretra, que va por el interior del pene en los machos, hasta
llegar al exterior. En las hembras la uretra se abre al vestíbulo, bien sea en
el ápice del clítoris, como en las ratas, o en cualquier otro punto.
Enfermedades
Del Sistema Excretor
El sistema excretor como los
demás sistemas que conforman a un organismo puede ser severamente afectado por
una serie de enfermedades, producidas por factores genéticos (a causa de
mutaciones causadas por agentes químicos como pesticidas y otros), factores
prenatales y por factores alimenticios en individuos adultos.
Las afecciones más comunes
son los casos en los cuales uno de los dos riñones aparece sin su
correspondiente uréter de tal forma que un riñón no es funcional para este
individuo.
Otro caso es cuando el individuo
nace únicamente con un riñón, este presenta dos uréteres y es en ocasiones
incapaz de cumplir la función renal en este individuo.
Se han registrado casos en
los cuales los fetos nacen con sus dos riñones pero estos no presentan uréteres
y esto causa la muerte del feto.
En las personas adultas el
riñón se lesiona con mucha frecuencia por malos hábitos alimenticios, por el
consumo desproporcionado de alcohol y por otros factores, causando en ocasiones
cálculos renales, cáncer de riñón y otras enfermedades que pueden llevar en
muchos casos a la extracción del órgano afectado.
Tomado de: http://www.monografias.com/trabajos5/sisex/sisex.shtml#rena
martes, 23 de septiembre de 2014
martes, 16 de septiembre de 2014
Métodos de Colecta Entomológica - Taller: El Mundo de los Insectos.
Colecta y Conservación de los Insectos
Materiales y Métodos de colecta
Para la colecta de insecto se han fabricados equipos, que son esenciales para la obtención de material entomológico. Los Métodos de colecta están basados sobre una gran cantidad de técnicas que se han ido perfeccionando con el tiempo.
Materiales de colecta.
A.-Red entomológica o Jama, esta puede ser:

Red Aérea: esta es una red de un tamaño considerable, que puede ser de 1,50 hasta 6 mts. La cual es confeccionada de materiales livianos (Aluminio, alambres acerados y tul).
Red de Rastreo: Esta es más corta que la anterior la cual es recomendable desde 1.20 cms. La cual debe ser de un tul más grueso y de mejor calidad.
Red Acuática: se pueden utilizar las anteriores, pero las hay especiales.
b.-Paraguas: Es un instrumento esencial para la obtención de muestras. Este consiste en una tela (Blanca) cuadrada (También hay circulares), la cual es sostenida por dos varillas en cruz (Estas pueden ser de aluminio).

c.-Aspirador: Es un aparato que presta mucha utilidad, principalmente para insectos muy pequeños. Este esta formado por un tubo (Plástico o de vidrio) y dos mangueras (Pueden ser sondas), cuya función es succionar los insectos al interior de éste.

Dentro de los métodos de colecta podemos encontrar trampas como:
1.-Trampas Barber: Esta consiste en recipientes (vasos plásticos o tarros) enterrados a nivel del suelo.

2.-Embudo Berlese: Gran cantidad de insectos vive sobre la hojarasca, musgos, etc. Este consiste en un embudo con una recipiente para contener la materia vegetal separada por una rejilla metálica y en extremo del embudo un recipiente con alcohol.

Los insectos que se encuentra en la materia vegetal tienden a enterrarse deslizándose por el embudo al recipiente con alcohol. Para acelerar este proceso se recomienda en época de verano exponerlos a la luz solar, en caso contrario someterlos a luz artificial.
3.-Trampa de luz: Es un buen método, ya que muchos insectos son atraídos por la luz durante la noche. Esta puede ser de 2 tipos; ya sea, instalando una ampolleta sobre un recipiente con alcohol, o empleando una tela blanca extendida en el suelo (también se puede colgar la tela entre dos arboles formando una especie de J) y en su centro una lámpara este tipo de trampa es una de las mas efectivas, es aconsejable utilizarla en noches cálidas y lugares despejados.
Trampas de Luz

4.-Trampa Malaise: Esta trampa está diseñada principalmente para capturar insectos voladores, la trampa en una especie de carpa que tiene en su parte mas alta se instala un recipiente con alcohol, la efectividad de esta trampa depende de su ubicación , conviene instalarla en lugares por donde circulen los insectos voladores.
Es una trampa poco utilizada, debido a su costo, la cual se puede fabricar de tubos y tul. Esta trampa da buenos resultados.

jueves, 11 de septiembre de 2014
jueves, 4 de septiembre de 2014
viernes, 29 de agosto de 2014
lunes, 25 de agosto de 2014
Educación Ambiental - 7°
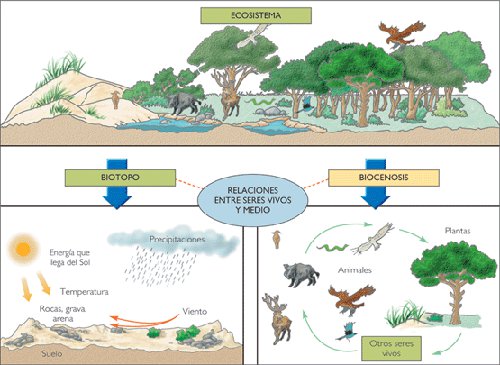
DINÁMICA DE LOS ECOSISTEMAS
1 - La dinámica de los ecosistemas es la parte de la ecología que estudia el funcionamiento de los ecosistemas, es decir, su evolución a lo largo del tiempo.
Dinámica de poblaciones
Es el estudio de los cambios que suceden en las poblaciones a medida que transcurre el tiempo, por abundancia o carencia de alimentos, condiciones climáticas, presencia de seres vivos competidores o depredadores, etc.
- Densidad: nº de individuos por unidad de espacio (superficie en los terrestres, volumen en los acuáticos). Depende de estos factores:
Tasa de natalidad: media de nacimientos en un año por cada mil habitantes (se expresa en tanto por mil)
Tasa de mortalidad: media de fallecimientos en un año por cada mil habitantes.
- Saldo migratorio: diferencia entre el nº de individuos que dejan la población y el nº de individuos que llegan a ella.
- Pirámides de edad: gráficos que representan la composición de una población en un momento determinado (edad, sexo).
- El crecimiento de una población es la variación de su tamaño a lo largo del tiempo. Presenta tres etapas: muy rápido al principio; cuando la población aumenta, el nº de nacimientos y muertes tiende a igualarse; y finalmente, la población se estabiliza.
- Factores limitantes del crecimiento ilimitado de las poblaciones (resistencia ambiental): limitantes externos (luz, agua, temperatura, enfermedades, contaminantes, depredadores, parásitos, etc.) y limitantes internos (canibalismo, huída, agresividad de las crías, etc.).
2 - Estabilidad y diversidad de un ecosistema
Diversidad: riqueza y número de especies que compone un ecosistema.
Estabilidad: cuando un ecosistema mantiene una serie de características que permiten reconocerlo a pesar de las perturbaciones.
Cuanto más diversos son los ecosistemas, más estables son (debido a la gran cantidad de relaciones). Y cuanto más diversos y estables son, mayor grado de organización presentan. Según el grado de organización, los ecosistemas son más o menos vulnerables a los cambios. Si el grado de organización es alto, el ecosistema se puede recuperar tras la explotación.
3 - Sucesión ecológica: son los cambios que se producen de forma natural en un ecosistema y que lo convierten en otro más ordenado y complejo.
4 - Comunidad clímax
Un ecosistema ha alcanzado el clímax cuando ha conseguido el mayor grado de estabilidad: no hay sustitución de unas especies por otras y la relación entre los seres vivos y el medio es muy estrecha.
ACCIÓN HUMANA SOBRE LOS ECOSISTEMAS
Relaciones entre el ser humano y el medio ambiente
Los seres humanos son capaces de modificar drásticamente el medio natural que les rodea, para adaptarlo a sus necesidades.
El ser humano ha llegado a crear un nuevo ecosistema: las grandes ciudades, con todos sus problemas y características.
El progreso ha requerido de grandes cantidades de combustible. Al principio sólo se utilizaba carbón vegetal, produciendo la deforestación de grandes zonas con el peligro de erosión y desertización.
Problemas ambientales derivados de la actuación humana
La actuación humana sobre el medio ambiente se traduce en una pérdida de biodiversidad o de variedad de formas de vida.
Extinción de especies: Las causas fundamentales que provocan la extinción de las especies son: la introducción de nuevas especies, la destrucción del hábitat y la caza y el exterminio deliberado.
Principales problemas ambientales
Desaparición de los grandes bosques y desertización
Si se suprimen árboles, el agua de la lluvia no es absorbida y corre por el suelo, y con frecuencia se lleva parte del suelo que ya no está fijado por las raíces.
Causas:
Explotación abusiva de los suelos pobres.
Pastoreo excesivo en pastos pobres.
Tala de árboles en las tierras secas.
Deforestación en las tierras altas.
Sistemas de riego inapropiados.
Contaminación del agua, el suelo y el aire
Los recursos naturalesson de tres tipos:
- Perennes: podemos disponer de ellos permanentemente: energía solar, mareas, viento...
- Renovables: pueden recuperarse, tras ser utilizados, mediante ciclos naturales haciendo posible que estén disponibles de forma más o menos constante: aire, agua, suelo, vegetales...
- No renovables: se consumen a mayor velocidad de la que se producen y, por lo tanto, se agotan: combustibles fósiles (carbón, petróleo) y minerales
Se considera que el agua, el aire o el suelo están contaminados cuando contienen sustancias, de origen natural o humano, en cantidades lo suficientemente grandes como para alterar su composición y provocar efectos indeseables.
El agua se contamina con las basuras que se arrojan a los ríos y mares y con el petróleo de algunos barcos que sufren accidente.
Los principales contaminantes del suelo son los fertilizantes (orgánicos e inorgánicos)y los plaguicidas.
La contaminación del aire hace que aumente la cantidad de ciertos gases y que disminuya la de otros, alterando el equilibrio de la atmósfera.
La lluvia ácida
Debida a la contaminación del aire por los humos que lanzan a la atmósfera algunas fábricas, los coches, las viviendas y las centrales térmicas. Estos humos están cargados de sustancias químicas que al combinarse con la humedad del aire forman ácidos que caen al suelo con la lluvia. Afecta sobre la salud del ser humano, destruye la vegetación, la vida en los ríos y lagos, e incluso deteriora los edificios.
El efecto invernadero
La Tierra se calienta gracias al calor que procede del Sol; la atmósfera retiene parte de ese calor y evita que se pierda en el espacio, contribuyendo a mantener caliente el planeta, actuando como un invernadero.
Si los gases de invernadero aumentan se produce un recalentamiento del planeta aumentando las temperaturas medias. El principal gas responsable es el CO2 .
El agujero de la capa de ozono
El ozono, que es un compuesto de oxígeno (03), forma una capa en la atmósfera que se encarga de filtrar los rayos ultravioleta enviados por el Sol.
Los gases clorofluorocarbonados o CFCs utilizados en neveras, aires acondicionados, sprays, etc. son los responsables de destruir esa capa de ozono y restar potencial para filtrar los rayos UVA con efectos indeseables como: irritación de ojos y aparato respiratorio, enfermedades cutáneas, nieblas tóxicas, etc.
Residuos tóxicos
Los residuos tóxicos pueden producir enfermedades o la muerte envenenando a los seres vivos bien por contacto directo, o bien a través de la cadena alimentaria. Para evitar esos problemas hay que aislar, enterrándolos, quemándolos o dejándolos en vertederos controlados, esos residuos.
Acciones de conservación del medio natural
Los principales objetivos de la conservación del medio natural son:
La preservación de los procesos básicos que mantienen la vida, y que son imprescindibles para la producción de alimentos, la salud, etc.
Mantener la diversidad de animales y plantas.
Asegurar el aprovechamiento sostenible de las especies y ecosistemas.
4.- Acciones de recuperación del medio natural
Reciclar
Reducir el consumo
Cambiar de hábitos
No contaminar
Tomado de:
http://www.cepaplusultra.edurioja.org/index.php/recursos/esecundaria/secundariadistancia/naturaleza/modulo-2/resumenes/117-dinamica-de-los-ecosistemas.html
Tomado de:
http://www.cepaplusultra.edurioja.org/index.php/recursos/esecundaria/secundariadistancia/naturaleza/modulo-2/resumenes/117-dinamica-de-los-ecosistemas.html
Equilibrio Dinámico de la Naturaleza.
Apesar de las constantes variaciones ambientales, los organismos tienden a mantener cierta estabilidad de las condiciones internas. Por ejemplo, si tomas tu temperatura corporal en días calurosos o muy fríos, te sorprenderás al ver que la temperatura será semejante a pesar de las grandes variaciones a las que estuviste sometido.
Un sistema vivo debe solucionar importantes problemas, tales como la regulación de la temperatura corporal, la cantidad de agua y sales, la concentración de azúcar sanguínea (glicemia) y la eliminación de productos de desecho, entre otras. Esto se logra mediante el funcionamiento coordinado de todos los tejidos y sistemas corporales, es decir la homeostasis. La demostración teórica de comportamiento caótico en poblaciones biológicas, de tal manera que las interacciones entre los componentes de un ecosistema pueden dar lugar a desestabilizaciones y extinciones (May, 1974), también ha contribuido a recelar de la idea de equilibrio.
Un sistema vivo debe solucionar importantes problemas, tales como la regulación de la temperatura corporal, la cantidad de agua y sales, la concentración de azúcar sanguínea (glicemia) y la eliminación de productos de desecho, entre otras. Esto se logra mediante el funcionamiento coordinado de todos los tejidos y sistemas corporales, es decir la homeostasis. La demostración teórica de comportamiento caótico en poblaciones biológicas, de tal manera que las interacciones entre los componentes de un ecosistema pueden dar lugar a desestabilizaciones y extinciones (May, 1974), también ha contribuido a recelar de la idea de equilibrio.
Los primeros intentos conc.eptuales para incorporar el hecho de que los sistemas naturales, a escala local, no se encuentran en equilibrio corresponden a los modelos de "dinámica de manchas" (patch dynamics), en los que las abundancias de las poblaciones, las interacciones entre especies y los ciclos de materia se consideran abiertos a la migración o flujo lateral (Pickett y White, 1985; Polis y Hurd, 1996).
Aunque la idea de tales modelos conceptuales vuelve a ser la de un equilibrio dinámico, denominado metaestabilidad" (Naveh, 1987; Wu y Loucks, 1995)
Según Nicolau (2006) Hay numerosos ejemplos en la naturaleza. El más claro es el denominado equilibrio químico, en el que varias sustancias coexisten en un sistema reaccionado entre sí pero manteniendo las proporciones de cada una de ellas. Cuando se alteran las condiciones en forma externa (por ejemplo aumentando la temperatura), favoreciendo la reacción en un sentido, se crea una nueva condición de equilibrio dinámico, con proporciones diferentes.
Cuando se incorpora al análisis los organismos vivos y su comportamiento, a simple vista parecería que el concepto de equilibrio dinámico es totalmente aplicable, pues en un ecosistema existen cambios internos (nacimientos, muertes, depredación, etc.) .
El paradigma del equilibrio en la naturaleza no sólo ha guiado a la ecología como ciencia, sino también al pensamiento social. Ante la idea del equilibrio en los sistemas naturales se han dado dos tipos de respuestas opuestas en nuestra sociedad (Terradas, 2001). La que trata de evitar o limitar al máximo la intervención humana sobre la naturaleza para no alterar dicho equilibrio (calificado de frágil).
Y la que considera que la naturaleza es capaz de soportar cualquier alteración antrópica dada su capacidad de recuperar el estado de equilibrio, por lo que no es exigente regulando las intervenciones humanas. La primera posición, minoritaria, está restringida a círculos ambientalistas extremos. La segunda, ha sido predominante durante la Revolución Industrial y ha guiado el consiguiente desarrollo económico. Uno de sus principales sustentos ideológicos es el elevado valor que concede a la tecnología, considerada como una herramienta con "poderes infinitos" para devolver a los sistemas natural y social al estado de equilibrio.
Esta percepción se encuentra actualmente cuestionada ante la evidencia de la existencia del Cambio Global, fenómeno que pone de manifiesto la ausencia de ese supuesto estado de equilibrio de la biosfera, así como la impredecibilidad de la respuesta de la misma. Ante el reconocimiento de que el paradigma del equilibrio no se cumple y de que las perturbaciones causadas por las actividades socioeconómicas está en el origen de los cambios que está experimentando la biosfera (deterioro ambiental), se ha elaborado un nuevo concepto:
El desarrollo sostenible.
El desarrollo sostenible tiene como objetivo disminuir la magnitud de las perturbaciones antrópicas y evitar así, que los parámetros bióticos y abióticos de la biosfera se salgan de los rangos de valores más propicios para el bienestar humano.
Tomado de:
http://datateca.unad.edu.co/contenidos/358006/contLinea/leccin_30_equilibrio_dinmico_de_la_naturaleza.html
lunes, 11 de agosto de 2014
Película: Gattaca (904)
Buen día, aquí está el link en donde pueden terminar de ver Gattaca:
Al ingresar a esa página deben hacer click sobre esta imagen:

Recuerden que deben enviar el trabajo escrito máximo el martes 18 de agosto a las 7:00 pm, a este correo:
sergioagx@yahoo.es
Hasta Pronto.
viernes, 8 de agosto de 2014
lunes, 28 de julio de 2014
Microscopio de Efecto Túnel - (Química 602 - 604)
¿Se pueden ver los átomos?
Por Paloma Fernández Sánchez
Actualmente hay muchas formas de ver o de medir "cosas” con resolución atómica, es decir de ver los átomos. En esta ocasión vamos a fijarnos en una de ellas: el Microscopio de Efecto Túnel. Este microscopio fue diseñado por Gerd Binning y Heinrich Röhrer y merecieron por ello el Premio Nobel en el año 1986. Con este pequeño artículo queremos rendir un homenaje a Heinrich Röhrer fallecido en mayo de este año.
El desarrollo del microscopio de efecto túnel y posteriormente de todos los agrupados bajo la categoría de Microscopias de Campo Cercano, han constituido una herramienta básica para el estudio de los materiales en los últimos años. Es evidente que muchos de los avances en el campo de la nanotecnología y la nanociencia no habrían sido posibles sin el desarrollo de este instrumento, pero no sólo esta rama se ha beneficiado de él. Aunque normalmente no nos demos cuenta de ello, la superficie de los materiales es un elemento fundamental, y en muchos procesos importantes juega un papel determinante. El microscopio de efecto túnel ha permitido no sólo saber dónde y cómo están colocados los átomos en la superficie, sino incluso moverlos.

Imágenes de los átomos de una superficie de óxido de titanio y de la construcción de una estructura que se conoce como corral cuántico, fabricada moviendo átomos de hierro sobre una superficie de cobalto.
En realidad el efecto túnel no es algo tan distinto de lo que conocemos de la vida diaria… ¡Bueno, un poco distinto sí!

Cuando viajamos por una zona montañosa, formada por muchas colinas y valles, si queremos ir de un valle a otro, no hay más remedio que subir la montaña y luego bajar otra vez. Pero eso es cansado, largo y a veces peligroso¡imagina bajar por la pendiente de la montaña de la imagen! Para hacer que los viajes sean más rápidos y cómodos se hacen túneles que nos permiten atravesar las montañas fácilmente con un coche u otro medio de transporte, o incluso andando. Pero hay túneles un poco especiales por los que no se mueven coches ni trenes, sino partículas como los electrones y que no están hechos para atravesar montañas, sino "capas de material”, o "espacios vacíos”.
Empecemos por el principio. Los materiales están hechos de átomos unidos unos a otros formando los materiales sólidos tal como los vemos, nosotros no distinguimos los átomos porque son muy pequeños, pero están ahí. Son tan pequeños que las leyes de la Naturaleza a las que estamos acostumbrados en nuestro mundo, dejan de funcionar y empiezan a pasar cosas nuevas o distintas.

Veamos un ejemplo. Imagina que tiras una pelota contra una pared, la pelota no puede atravesar la pared, rebotará y volverá a tus manos, a menos que la lances con suficiente energía como para que pueda saltar por encima.
Sin embargo si tirases la pelota contra una pared de un material frágil y no muy gordo o contra una ventana podría pasar otra cosa. Si tirásemos la pelota lo suficientemente fuerte podríamos atravesar la pared o ventana, aunque eso sí, rompiendo el cristal. ¡Pero si estuviéramos a escala atómica habríamos podido atravesar la pared sin romperla! Y a eso es a los que los físicos llaman "efecto túnel”. Eso sí la pelota tendría que ser un electrón y la pared una lonchita de material muy muy pequeña.
Lo que hemos llamado efecto túnel puede ocurrir también entre dos superficies metálicas separadas por una distancia muy pequeña. Normalmente, los electrones no podrán abandonar el material y saltar de una superficie a la otra, pero en algunas condiciones podrán salvar esa distancia, y aparecer en la otra superficie.
Imagina que estás en el terreno montañoso del dibujo y para salir tienes que saltar y agarrar las asas de la parte de arriba. El terreno montañoso de abajo representa la superficie del material, con las montañas en el papel de los átomos. El muñeco es el electrón que tiene que conseguir atravesar la distancia entre las montañas y los valles y la punta de arriba, donde están las asas. Si el muñeco está en el valle, aunque salte, no llegará a alcanzar las asas, pero si está en la parte alta seguramente sí podrá.

Si estuviéramos mirando al muñeco, cada vez que éste alcanzara las asas sabríamos que había saltado desde un punto alto, por lo tanto podríamos hacer un esquema de dónde están localizados los puntos más altos. Si la punta está enfrente de un valle, aunque el muñeco salte no alcanzará, si está en la cima de la montaña con un pequeño salto podrá alcanzar, y si está en un punto intermedio de la ladera, podrá alcanzar las asas pero con mayor esfuerzo, saltando más.
El microscopio de efecto túnel hace algo parecido. La punta se va moviendo muy cerca de la superficie que estamos estudiando, pero sin rozar. Cuando los electrones de la superficie sean capaces de atravesar el espacio entre la punta y a muestra, mediremos una corriente eléctrica, pero esto sólo ocurrirá cuando la distancia de separación entre la punta y la muestra sea lo suficientemente pequeña como para que se produzca el efecto túnel. Si mantenemos la altura de la punta constante, los cambios en la corriente que medimos estarán asociados a la topografía de la muestra. También podríamos fijar la corriente que deseamos medir y dejar que la aguja alcance la altura necesaria para que se produzca, en cualquiera de los dos casos obtendríamos un mapa de la topografía de la superficie. Si la aguja es suficientemente fina, seremos capaces de ver los átomos, ya que cuando la punta esté justo enfrente de un átomo de la superficie será cuando pueda producirse el efecto túnel de manera más eficiente, es decir tendremos una corriente máxima. De esta forma podemos ver la superficie de las cosas con gran detalle, en realidad hablamos de resolución atómica, o sea que podemos distinguir los átomos que forman la superficie. En el esquema de la parte derecha de la figura puedes ver a qué nos referimos.

Esquema básico de un microscopio de efecto túnel.
Mira cómo se parecen los dos dibujos

Tomado de: http://enciende.cosce.org/boletin/index.asp?item=78
martes, 15 de julio de 2014
lunes, 14 de julio de 2014
miércoles, 28 de mayo de 2014
Pelicula: La Isla - 604
En el siguiente Link pueden terminar de ver la película:
sábado, 17 de mayo de 2014
martes, 13 de mayo de 2014
Moléculas - Química 601 - 603
Tomado de: http://www.slideshare.net/profesorleonardo/qumica-para-nios-01
Suscribirse a:
Entradas (Atom)